

Abstract
Information processing in the retina starts at the first synaptic layer, where photoreceptors and second-order neurons exhibit a complex architecture of glutamatergic and electrical synapses. To investigate the composition of this highly organized synaptic network, we determined the spatial relationship of zonula occludens-1 (ZO-1) with different connexins (Cx) and glutamate receptor (GluR) subunits in the outer plexiform layer (OPL) of rabbit, mouse, and monkey retinas. ZO-1 is well known as an intracellular component of tight and adherens junctions, but also interacts with various connexins at gap junctions. We found ZO-1 closely associated with Cx50 on dendrites of A-type horizontal cells in rabbit, and with Cx57 at dendro-dendritic gap junctions of mouse horizontal cells. The spatial arrangement of ZO-1 at the giant gap-junctional plaques in rabbit was particularly striking. ZO-1 formed a clear margin around the large Cx50 plaques instead of being colocalized with the connexin staining. Our finding suggests the involvement of ZO-1 in the composition of tight or adherens junctions around gap-junctional plaques instead of interacting with connexins directly. Furthermore, gap junctions were found to be clustered in close proximity to GluRs at the level of desmosome-like junctions, where horizontal cell dendrites converge before invaginating the cone pedicle. Based on this distinct spatial organization of gap junctions and GluRs, it is tempting to speculate that glutamate released from the photoreceptors may play a role in modulating the conductance of electrical synapses in the OPL.
Introduction
Retinal processing begins in the outer plexiform layer, where cone pedicles and rod spherules provide multiple output synapses onto bipolar and horizontal cells. Synaptic transmission is mediated by glutamate and metabotropic or ionotropic glutamate receptors (GluRs) (for review, see Wässle, 2004), and by electrical coupling of different cell types via connexins (for review, see Söhl et al., 2005). These components are accompanied by different scaffolding proteins and further cell–cell contacts to stabilize the synaptic organization of the neuronal network.
Some of the proteins involved in these complex synapses and their location in the OPL have already been analyzed, but the question regarding their spatial relations to each other remains open. To address this question and to analyze whether the synaptic architecture in the OPL is conserved between mammalian species, we investigated the expression patterns of the scaffolding protein zonula occludens-1 (ZO-1), different connexins (Cx), and GluRs on horizontal cells of rabbit, mouse, and monkey retinas.
Horizontal cells are homologously coupled by gap junctions, forming an extensive network that feeds back onto the photoreceptors. Because of this electrical coupling, each horizontal cell receives photoreceptor input over a receptive field that is much larger than its dendritic field (Kaneko, 1971), and the spatial extent of the feedback signal is determined, in part, by the strength of coupling in the horizontal cell network (Dacheux and Raviola, 1982; Bloomfield et al., 1995). Horizontal cell coupling is regulated by ambient light (Xin and Bloomfield, 1999) via neuromodulators such as dopamine and retinoic acid (Teranishi et al., 1983; Perlman and Ammermüller, 1994; He et al., 2000; Pottek and Weiler, 2000). Horizontal cells in mouse are coupled by gap junctions composed of Cx57 (Hombach et al., 2004; Shelley et al., 2006; Janssen-Bienhold et al., 2009); A-type horizontal cells in rabbit retina are coupled by Cx50 (O'Brien et al., 2006). Furthermore, numerous proteins have been identified that may be involved in the mechanism of gap-junctional coupling. For instance, ZO-1 has been found at gap junctions, interacting with several different connexins (for review, see Giepmans, 2004). However, the structural role of ZO-1 is unclear. Studies in mouse retina revealed an association of ZO-1 with Cx36 in both the OPL and the inner plexiform layer (IPL), and with Cx45 in the IPL (Li et al., 2004, 2008; Ciolofan et al., 2006, 2007).
In the present study, we found that ZO-1 was colocalized with Cx36 at rod spherules and at the cone pedicle base in all three species. However, ZO-1 was also clustered in close proximity to GluRs at desmosome-like junctions between horizontal cell dendrites. Here, ZO-1 was colocalized with Cx50 on A-type horizontal cells in rabbit, and with Cx57 at dendro-dendritic gap junctions of mouse horizontal cells. GluRs at the desmosome-like junctions 1–2 μm below the cone pedicles are thought to be activated by glutamate released from the cone pedicle, with each side of the junction corresponding to a postsynaptic density (Haverkamp et al., 2000).
We also studied the spatial relationship of Cx50 and ZO-1 at giant gap-junctional plaques of A-type horizontal cells in rabbit and found that ZO-1 encloses the Cx50 plaques, rather than directly colocalizing with the connexin staining.
Materials and Methods
Animals and tissue preparation
Wild-type mice (C57BL/6J) were deeply anesthetized with isoflurane and decapitated before tissue dissection. Macaque monkeys (Macaca fascicularis) and rabbits (Chinchilla Bastard) were killed by an overdose of pentobarbitone (100 mg/kg) given intravenously in experiments not related to the present study, and the eyes were enucleated postmortem. All procedures were approved by the local animal care committee and were in accordance with the law for animal experiments issued by the German government (Tierschutzgesetz).
The posterior eyecups were immersion fixed in 2 or 4% paraformaldehyde (PFA) in 0.1 m phosphate buffer (PB), pH 7.4, for 10–30 min at room temperature. Following fixation, retinas were dissected from the eyecup, cryoprotected in graded sucrose solutions (10, 20, 30% w/v), and stored at −20°C in 30% sucrose until use. Retinal pieces were either used as a whole mount or sectioned vertically at 16 μm using a cryostat.
Antibodies and light microscopic immunocytochemistry
The primary antibodies used in the present study are listed in Table 1.
Primary antibodies used in this study
Immunocytochemical labeling was performed using the indirect fluorescence method. Sections were incubated overnight with primary antibodies in 5% normal donkey serum (NDS), 1% bovine serum albumin (BSA), 0.5% Triton X-100, and 0.02% sodium azide in PB. After washing in PB, secondary antibodies were applied for 1 h. These were conjugated either to Cy3 (Dianova), or Alexa TM 488 (Invitrogen). Retinal pieces were incubated freely floating for 2 d in a mixture of primary antibodies containing 5% NDS, 1% BSA, and 1% Triton X-100, followed by a 3 h incubation in a mixture of secondary antibodies.
Confocal photomicrographs were taken using a Zeiss LSM 5 Pascal confocal microscope equipped with an argon and an HeNe laser. High-resolution scanning of image stacks was performed with a Plan-Apochromat 63×/1.40 oil-immersion objective at 1024 × 1024 pixels and a z-axis increment of 0.32 μm. When projections of image stacks are shown, 3–5 serial optical sections were collapsed into a single plane. Brightness and contrast of the final images were adjusted using Adobe Photoshop CS v8.0.1.
Microinjections
Isolated pieces of retinas of C57BL/6J mice were incubated in Ames solution (pH 7.4, Sigma) containing 10 μm DAPI (Sigma) for 60 min at room temperature and mounted on filter paper (Millipore) with ganglion cell layer up.
Horizontal cells were microinjected with 2% Lucifer yellow (Sigma) and 4% Neurobiotin (Vector Laboratories) as previously described by Shelley et al. (2006). After injection, pieces were fixed in 4% PFA in PB for 15 min. To visualize the Neurobiotin, the tissue was incubated with Cy3-conjugated streptavidin (Dianova) overnight in PB and 1% Triton X-100 at 4°C. Subsequently, the retinal pieces were labeled with antibodies against ZO-1, following the protocol as described above.
Preembedding immunoelectron microscopy
Preembedding immunoelectron microscopy was carried out on vibratome sections as described in detail previously (Sassoè-Pognetto et al., 1994). After preincubation in PBS containing 10% normal goat serum (NGS) and 1% BSA, vibratome sections were incubated for 4 d at 4°C in primary antibodies diluted in 3% NGS and 1% BSA in PBS.
Postembedding immunoelectron microscopy
The slam-freezing and cryosubstitution methods used in this study are a modification of those previously described (Baude et al., 1993). Briefly, small pieces of retina were slam-frozen onto a precooled copper block using a Reichert MM80 unit (Leica). The frozen specimens were then transferred to a cryosubstitution unit (Reichert AFS; Leica) and placed into a solution of 0.5% uranyl acetate (w/v) in 100% methanol at −90°C. After 36 h, the temperature was increased stepwise to −30°C. The samples were then washed several times in precooled methanol and then progressively infiltrated with Lowicryl HM20 (Chemische Werke Lowi; 1:1 Lowicryl to methanol, 90 min; 2:1 Lowicryl to methanol, 90 min; 100% Lowicryl, 90 min; 100% Lowicryl, overnight). Polymerization was achieved by exposure to UV light for 12 h.
Ultrathin sections were cut and collected on Formvar-coated nickel grids. The sections were first incubated in 0.05 m Tris-buffered saline (TBS), pH 7.6, for 15 min, followed by a blocking solution containing 10% NGS and TBS with 0.1% Triton (TBST) for 20 min. Sections were then incubated overnight in primary anti-GluR2/3 diluted 1:10 or anti-GluR4 diluted 1:5 in 1% NGS and TBST. After washing in TBS, sections were incubated for 2 h in secondary anti-rabbit IgG conjugated to 10 nm gold (ICN Biomedicals) diluted 1:20 in 1% NGS and TBST. After further washing in TBS and PB, the sections were fixed for 5 min in 2% glutaraldehyde in PB, washed in distilled water, and counterstained with uranyl acetate and lead citrate. The grids were then viewed with a Zeiss EM10 electron microscope.
Results
Zonula occludens-1 expression in the mammalian retina
The overall ZO-1 expression pattern in rabbit and macaque monkey retina (Fig. 1) appeared very similar to that seen in the mouse retina (Li et al., 2004; Ciolofan et al., 2006, 2007) (Fig. 1A). We found strong ZO-1 immunoreactivity at the outer limiting membrane (OLM), and punctate staining in the OPL and IPL. Furthermore, the tight junctions between endothelial cells of the blood vessels were labeled in mouse and monkey retina, and also some amacrine cell bodies in the inner nuclear layer (INL) showed immunoreactivity. In these animals, the ZO-1-immunoreactive puncta in the OPL were densely clustered in regular arrays, most likely below the pedicles of cone photoreceptors. In rabbit retina, the clustering at cone pedicles was not as obvious as in the two other species, but large plaques of ZO-1 immunoreactivity were observed in the proximal part of the OPL. The plaques in the rabbit retina pointed to an expression of ZO-1 by horizontal cells, whose dendrites are present at those sites in the OPL.

Expression pattern of ZO-1 in the mammalian retina. A–C show projections of image stacks of vertical cryostat sections labeled with antibodies against ZO-1 in mouse (A), rabbit (B), and monkey (C) retinas. Retinal ZO-1 expression is similar among the three species: the outer limiting membrane (OLM) is strongly labeled and punctate ZO-1 staining is found throughout the outer and inner plexiform layers (OPL, IPL). While the whole IPL is labeled homogeneously, ZO-1 in the OPL is also densely clustered in regular arrays most likely at cone pedicles; this is particularly apparent in the monkey retina. In the inner nuclear layer (INL) and the ganglion cell layer (GCL), ZO-1 is located at tight junctions between vascular endothelial cells (arrows). Furthermore, cell bodies of amacrine cells were labeled in the INL of monkey and mouse retinas (ONL, outer nuclear layer). Scale bars: 20 μm.
ZO-1 forms margins around gap junctions of A-type horizontal cells in rabbit
In rabbit retina, as in many mammalian species, there are two types of horizontal cells named A-type and B-type cells. A-type horizontal cells are axonless with large asymmetric dendrites that contact cones. B-type cells have numerous, symmetrical dendrites that also contact cones, and a long axon that expands into an elaborate axon terminal which selectively contacts rods (Bloomfield and Miller, 1982; Dacheux and Raviola, 1982). In the OPL of the rabbit retina, large ZO-1-immunoreactive plaques and smaller puncta were colocalized with A-type horizontal cells at the sites where their dendrites cross (Fig. 2A,B). A-type horizontal cells were labeled with antibodies against the calcium-binding protein calbindin (CaBP) (Röhrenbeck et al., 1987). Interestingly, the staining pattern of ZO-1 and CaBP in the proximal part of the rabbit OPL, as shown here, closely resembles the staining pattern of Cx50 and CaBP as shown recently in the rabbit retina by O'Brien et al. (2006) (their Fig. 4), suggesting that ZO-1 may interact with Cx50 in these cells.

ZO-1 is expressed by A-type horizontal cells (HCs) in rabbit retina and forms margins around Cx50-containing plaques. A, B, Whole-mounted rabbit retina labeled with antibodies against ZO-1 and calbindin (CaBP), which stains A-type HCs. ZO-1 immunoreactivity appears as punctate staining and large plaques (arrows). C–H, Whole-mounted rabbit retina immunostained for ZO-1 and Cx50. C–E show a large ZO-1-immunoreactive plaque as shown in B. F–H, The spatial arrangement of connexin clusters surrounded by ZO-1 also occurs at smaller gap junctions. Scale bars: (in B) A, B, 25 μm, (in H) C–H, 5 μm.
Figure 2C–E shows a gap junction in the OPL of a rabbit retina double labeled for ZO-1 and Cx50. This staining confirmed an association of the two proteins at the gap junctions between A-type horizontal cell dendrites. Furthermore, the huge size of these junctions allowed a good resolution of the spatial relationship between the labeled connexins and ZO-1. In fact, only a fraction of Cx50- and ZO-1-immunoreactive staining was directly colocalized. ZO-1 appeared instead to form margins around the connexin staining and to cross the gap junction with a grid-like structure as soon as the gap junction exceeds a certain size. The spatial arrangement of connexin clusters surrounded by ZO-1 staining could also be observed at smaller plaques (Fig. 2F–H).
Spatial relationship of ZO-1 to Cx50 and Cx57 on horizontal cells of rabbit and mouse
We compared rabbit and mouse retinas to investigate ZO-1 and its possible association with gap junctions beneath the cone pedicles. In the rabbit retina, Cx50 expressed by A-type horizontal cells was reported to cluster at the cone pedicles [O'Brien et al. (2006), their Fig. 11G]. We confirmed this finding on vertical sections of the rabbit retina (Fig. 3A,B) and could show, in whole-mounted specimen, that all Cx50-immunoreactive puncta were colocalized with the clustered ZO-1 at the cone pedicles (Fig. 3C–E). The colocalization of the ZO-1 clusters with Cx50 on the horizontal cell dendrites occurred at a distance of ∼1–2 μm from the cone pedicle base. This is the site where the horizontal cell dendrites converge before invaginating the cone pedicle. Most, if not all, of the ZO-1-immunoreactive puncta which were not colocalized with Cx50 were located at Cx36-containing gap junctions (see below; supplemental Fig. 1D–F, available at www.jneurosci.org as supplemental material). However, we cannot exclude that the unidentified connexin of the B-type horizontal cell dendrites is also associated with ZO-1 beneath cone pedicles.

Cx50 and ZO-1 are colocalized beneath cone pedicles in rabbit retina. A, B, Vertical cryostat section double labeled for Cx50 and the kainate receptor subunit GluR5. GluR5 is expressed on the tips of OFF bipolar cells and was used to label the base of the cone pedicles. Cx50 is loosely clustered beneath the cone pedicles. In addition, the Cx50 antibody faintly labels some somata in the INL which appear partially at the lower edge of the figure. C–E, Whole-mounted rabbit retina immunostained for ZO-1 and Cx50. At a level slightly distal to the large ZO-1 plaques, ZO-1 staining appears more punctate and sometimes clustered. The merged image of ZO-1 and Cx50 staining in E shows that all Cx50 puncta are colocalized with ZO-1 and that ZO-1 is clustered with Cx50 at the cone pedicles (circled). Scale bars: 5 μm.
The mouse has only one type of horizontal cell, of the axon-bearing B-type morphology (Suzuki and Pinto, 1986; Peichl and González-Soriano, 1994). Figure 4A–C shows the localization of ZO-1 immunoreactivity on the dendrites of a Neurobiotin-injected horizontal cell network. Like in rabbit, ZO-1 clusters were colocalized with horizontal cell dendrites beneath the cone pedicle base.

ZO-1 is expressed on horizontal cell (HC) dendrites in mouse retina and colocalized with Cx57 at dendritic sites. It is not located on HC axon terminals in mouse and rabbit retinas. A–C, Whole-mounted mouse retina labeled for ZO-1 after microinjection of Neurobiotin into a HC. The images show an optical section through the OPL slightly beneath the dendritic tips of the HCs, where the ZO-1 puncta beneath two cone pedicles are in focus. D–F, Double labeling of the photoreceptor terminals with PSD-95 and Cx57 in vertical sections of the mouse retina. The bases of two cone pedicles are marked by dashed lines in D and F. G–I, Vertical sections of mouse retina immunostained for Cx57 and ZO-1. Three putative pedicles can be identified and are marked by dashed lines. J–L, Double labeling of Cx57 and ZO-1 in the rabbit retina. Cx57 staining is only found above the cone pedicles (dashed lines). Scale bars: (in C) A–C, (in L) D–L, 2 μm.
Recently, it was shown that Cx57 is responsible for the electrical coupling of mouse horizontal cells (Hombach et al., 2004; Shelley et al., 2006) and Ciolofan et al. (2007) investigated the association of ZO-1, Cx36, and Cx57 in the mouse OPL. Beneath cone pedicles, they found densely clustered ZO-1 but no Cx57 expression, and hence no overlap between ZO-1 and Cx57. However, Janssen-Bienhold et al. (2009) found Cx57 expression on both dendrites (dendro-dendritic gap junctions) and axon terminals (axo-axonal gap junctions) of mouse horizontal cells. Therefore, we reinvestigated the expression of Cx57 and its association with ZO-1 in the mouse OPL. We used PSD-95 as a marker for photoreceptor terminals (Koulen et al., 1998). Double-labeling experiments with Cx57 and PSD-95 on vertical cryosections revealed Cx57-immunoreactive puncta clearly localized postsynaptic to rod spherules and cone pedicles (Fig. 4D–F). All Cx57-immunoreactive clusters found beneath cone pedicles (dendro-dendritic gap junctions) were exclusively colocalized with ZO-1, whereas Cx57 immunostaining beneath rods in the distal OPL (axo-axonal gap junctions) was not associated with ZO-1 (Fig. 4G–I).
ZO-1-immunoreactive puncta at the level of rod spherules were located at Cx36-containing gap junctions. The same holds true for the ZO-1 puncta beneath the cone pedicles, which were not colocalized with Cx57 (supplemental Fig. 1A–C, available at www.jneurosci.org as supplemental material). It has been shown previously that ZO-1 colocalizes with Cx36 in the OPL of the mouse retina (Ciolofan et al., 2007). Here, Cx36 forms gap junctions between dendritic tips of OFF cone bipolar cells and between photoreceptors (for review, see Söhl et al., 2005). We confirmed the finding of Ciolofan et al. (2007) in the mouse retina, and double labeling of ZO-1 and Cx36 in rabbit (supplemental Fig. 1D–F, available at www.jneurosci.org as supplemental material) and monkey retinas (see below; Fig. 6J–L) yielded the same results.
In the rabbit retina, Cx57 is expressed on the axon terminals of B-type horizontal cells (Pan et al., 2007). As expected, ZO-1 was not colocalized with Cx57 (Fig. 4J–L), which is comparable to the noncolocalization of ZO-1 and Cx57 in the distal part of the mouse OPL.
ZO-1 localization at cone pedicles of the macaque monkey retina
We used the monkey retina to investigate more precisely the expression pattern of the clustered ZO-1 in the OPL, and counterstained ZO-1 with antibodies labeling different cell types and synaptic components of gap junctions and glutamatergic synapses.
In the monkey retina, there are also two types of horizontal cells, H1 and H2 cells. The dendritic tips of H1 cells contact medium- and long-wavelength-sensitive (M- and L-) cone pedicles, while short-wavelength-sensitive (S-) cone pedicles are only sparsely contacted. The dendritic tips of H2 cells preferentially contact S-cone pedicles but have additional contacts with all cones within their dendritic field (Dacey et al., 1996; Goodchild et al., 1996). Both horizontal cell types have axons: the H1 cell axon terminal contacts rods exclusively and the H2 axon terminal has only sparse contacts to cone pedicles (Kolb et al., 1980). Figure 5A–C shows the ZO-1 staining in the OPL of the monkey retina, counterstained with antibodies against S-opsin. The ZO-1 staining in the whole-mounted retina revealed string-like plaques and further immunoreactive puncta between the clustered regions at putative cone pedicles (Fig. 5A). At the location of the S-cone pedicle, ZO-1 expression was strongly reduced compared with M- and L-cone pedicles, which indicated expression of ZO-1 by H1 cells. In addition to the specific labeling of S-cones, the JH455 S-opsin antibody cross-reacted with filaments of the cytoskeleton in dendrites and cell bodies of H1 horizontal cells in the macaque monkey retina (Fig. 5D–F). However, the cross-reactivity of this antibody, together with the ZO-1 staining, show that all string-like ZO-1 plaques were confined to dendrites of the H1 horizontal cell.

ZO-1 is expressed by H1 horizontal cells (HCs) in monkey retina. A–C, Whole-mounted macaque monkey retina immunostained for ZO-1 and S-opsin with the JH455 S-opsin antibody. One short-wavelength-sensitive (S)-cone pedicle is labeled by S-opsin (circled). ZO-1 staining is strongly reduced at the S-cone pedicle compared with the dense clustering at medium- and long-wavelength-sensitive cone pedicles. The JH455 antibody also cross-reacts with cytoskeletal filaments of dendrites and cell bodies (asterisks) of H1 HCs. All string-like ZO-1 immunoreactivity (arrowheads) is confined to H1 dendrites. D–F, Vertical cryostat section double labeled for calbindin (CaBP) and JH455 anti-S-opsin. D, CaBP labels cone photoreceptors, bipolar cells, and H2 cells in the outer monkey retina. Two labeled H2 cell bodies are shown. E, Besides its specific staining of short-wavelength-sensitive cones, the antibody against S-opsin cross-reacts with cellular structures of HC cell bodies and dendrites. The image shows dense staining in the OPL and three large cell bodies at the distal border of the INL. F, None of the S-opsin-immunoreactive HCs are positive for CaBP. Their overall morphology, the larger diameter of their cell bodies compared with H2 cells, and the fact that they are negative for CaBP provide good evidence that the S-opsin antibody labels H1 cells. Scale bars: (in C) A–C, 5 μm, (in F) D–F, 10 μm.
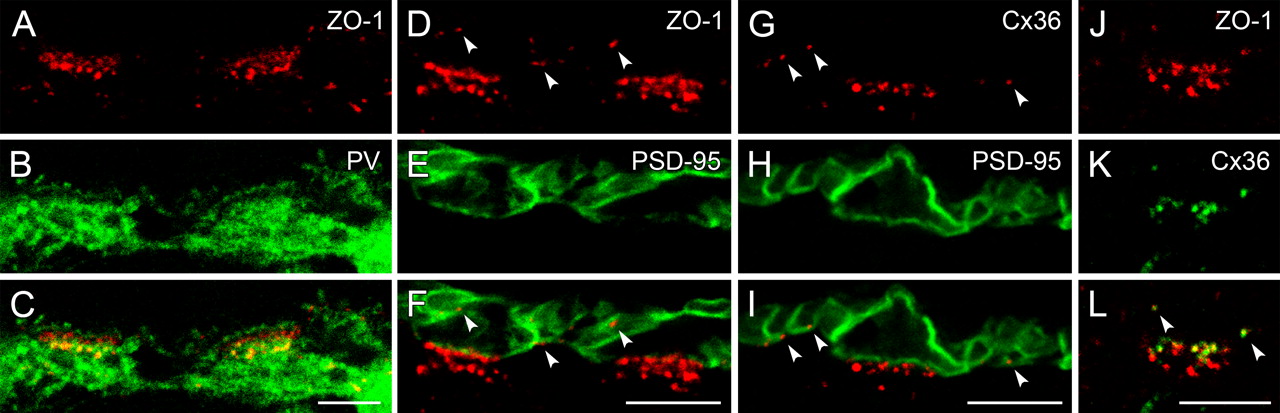
ZO-1 was clustered as two distinct bands at the cone pedicle base of the monkey retina, and further immunoreactive puncta appeared in more distal and proximal parts of the OPL (Fig. 6A–F). As we had observed in the other species, the lower ZO-1 band was colocalized with horizontal cell dendrites (Fig. 6A–C), where they form desmosome-like junctions with each other and where different ionotropic GluR subunits have been identified as postsynaptic elements of chemical synapses (Haverkamp et al., 2000). The upper band of ZO-1-immunoreactive clusters, which was located just above the level of the desmosome-like junctions, was not associated with horizontal cell processes. The remaining ZO-1 puncta in the proximal OPL were localized on either horizontal cell dendrites or cell bodies. Further puncta in the distal OPL were found slightly beneath the invaginating tips of horizontal cell axon terminals. Double labeling of ZO-1 and PSD-95 revealed that the upper band of ZO-1 coincided with the cone pedicle base (Fig. 6D–F), whereas the puncta in the distal OPL were all associated with rod spherules (arrowheads). To find out whether ZO-1 at the pedicle base and at the level of rod spherules is associated with Cx36-containing gap junctions, as in mouse and rabbit, we first analyzed the expression pattern of Cx36 in the primate retina (Fig. 6G–I). As expected, Cx36-immunoreactive puncta were found at the cone pedicle base and associated with rod spherules. Double-labeling experiments with ZO-1 and Cx36 supported the results from mouse and rabbit retinas: the upper band of ZO-1 at the pedicle base and the puncta associated with the membranes of rod spherules were all colocalized with Cx36 (Fig. 6J–L).

ZO-1 colocalizes with Cx36 in the OPL of monkey retina. A–C, Single optical slice of a vertical section immunostained for ZO-1 and parvalbumin (PV), which labels all horizontal cells (HCs). ZO-1 is clustered in two bands at two cone pedicles each. The lower bands are colocalized with PV at the level of the desmosome-like junctions, while the upper bands are situated between the desmosome-like junctions and the invaginating tips of HC dendrites. D–F, Projection of an image stack of vertically sectioned retina stained for ZO-1 and the photoreceptor terminals, labeled by antibodies against the postsynaptic density protein PSD-95. Two cone pedicles and the surrounding rod spherules are shown. The arrowheads point to ZO-1 puncta in the distal OPL, which are all closely associated with the membranes of rod spherules. G–I, Double labeling of Cx36 and PSD-95. Cx36 is expressed at the cone pedicle base and at rod spherules (arrowheads). J–L, The immunostaining of ZO-1 and Cx36 at a single cone pedicle shows that they are colocalized at the pedicle base and at rod spherules (arrowheads). Scale bars: 5 μm.
The connexins of primate horizontal cells are not yet known and therefore equivalent double-labeling experiments with ZO-1, as presented before in mouse and rabbit, were not feasible. To rule out a function of ZO-1 at the level of the desmosome-like junctions in the primate retina other than an association with gap junctions, we performed double-staining experiments of ZO-1 and different GluR subunits. Although ZO-1 expression has now been shown on horizontal cells at the same level with the desmosome-like junctions, it was not colocalized with the AMPA receptor subunits GluR2/3 (Fig. 7A–C). GluR2/3 and GluR4 are expressed at the dendritic tips of horizontal cells and at the desmosome-like junctions (Haverkamp et al., 2001a). In retinal whole mounts, ZO-1 and GluR2/3 clusters were found in close apposition (Fig. 7D–F). The same results were obtained by double labeling ZO-1 and GluR4 (Fig. 7G–H). The kainate receptor subunits GluR6/7 are also expressed at the desmosome-like junctions of horizontal cells, and in addition, on dendritic tips of OFF bipolar cells at the cone pedicle base (Haverkamp et al., 2001b). The results of double-labeling experiments with ZO-1 and GluR6/7 were similar to those shown with GluR2/3 and GluR4. The large ZO-1 puncta were not colocalized, but closely associated with GluR6/7 at the level of desmosome-like junctions (Fig. 7J–L). Since in rabbit retina, clusters of GluR2/3 and GluR4 subunits were also found beneath the cone pedicles (Pan and Massey, 2007), a close association to Cx50/ZO-1 was very likely, and was confirmed in double-labeled material with GluR2/3 and ZO-1 (data not shown). In summary, ZO-1 and the GluR subunits at the desmosome-like junctions were not randomly arrayed, but closely associated.

ZO-1 at the level of the desmosome-like junctions is closely associated with GluRs in monkey retina. A–C, Vertical cryostat section double labeled for ZO-1 and the AMPA receptor subunits GluR2/3. A single cone pedicle is shown, at which the GluR2/3 staining occurs as two bands. The upper band is located at the tips of invaginating horizontal cell dendrites, the lower one at the desmosome-like junctions. D–I, Horizontal view of the desmosome-like junctions labeled for ZO-1, GluR2/3 (D–F), and GluR4 (G–I) beneath cone pedicles in whole-mounted retina. Instead of a random distribution, the large ZO-1 puncta always appear in very close vicinity to the GluR puncta. J–L, Immunostaining of ZO-1 and the kainate receptor subunits GluR6/7 at the level of the desmosome-like junctions of two cone pedicles. Most of the ZO-1 puncta are closely associated with GluR6/7. Scale bars: 5 μm.
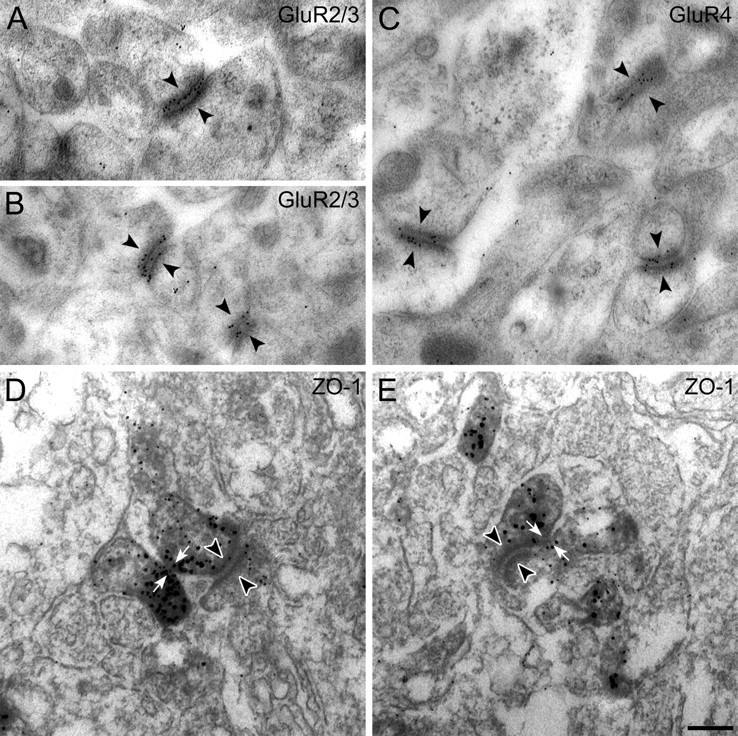
To further investigate the close association of the GluR subunits with ZO-1, we did preembedding and postembedding immunocytochemistry and electron microscopy. Figure 8A–C shows results of the localization of GluR2/3 and GluR4 at the desmosome-like junctions of macaque retina, using postembedding immunoelectron microscopy. Both sides of the junctions were often decorated with gold particles, suggesting that AMPA receptors are expressed by both members of this junction. These results show that the AMPA receptors are not distributed all over the two postsynaptic processes, as one might deduce from preembedding immunolabeling (Haverkamp et al., 2001), but are aggregated directly at the desmosome-like structure. Unfortunately, we were not able to produce ZO-1 postembedding immunolabeling. However, our serially sectioned preembedding material showed that the center zone of immunoreactivity at these sites was often associated with the contiguous plasma-membrane and not directly with the desmosome-like structure itself. The two examples in Figure 8, D and E, show putative gap junctions (white arrows) adjacent to desmosome-like junctions (arrowheads).

Electron micrographs of vertical sections through the synaptic complex beneath cone pedicles of the macaque monkey retina that were immunolabeled using postembedding (A–C) and preembedding methods (D, E). A, B, GluR2/3 label is located at both sides of the desmosome-like junctions (arrowheads) beneath the cone pedicle. C, Both sides of three desmosome-like junctions are labeled for GluR4 (arrowheads). D, E, Preembedding immunolabeling of ZO-1 beneath the cone pedicle. The white arrows point to putative gap junctions directly adjacent to desmosome-like junctions (arrowheads). Scale bar: 0.2 μm.
Discussion
This study describes the spatial relationship of ZO-1 to different connexins and glutamate receptors in the OPL of rabbit, mouse, and monkey retina. Our data show that the spatial organization of these proteins beneath the cone pedicles is very similar among mammalian species (Fig. 9A). Cx36 is expressed between photoreceptors and between dendrites of OFF bipolar cells (Feigenspan et al., 2004). GluRs are expressed in close association to gap junctions at the desmosome-like junctions. Additionally, the expression of ZO-1 in horizontal cells and the association with Cx50 or Cx57 is cell type specific: In monkey, ZO-1 is clustered beneath cone pedicles on H1 horizontal cell dendrites, but not on H2 cells. So far, 21 connexin genes have been described in the human genome (Söhl and Willecke, 2003); however, except for Cx36 (Massey, 2008) no other connexin protein has been detected in monkey retina to date. In rabbit, ZO-1 is associated with Cx50 on A-type horizontal cell dendrites. B-type cells express Cx57 on their axon terminals (Pan et al., 2007), which is not associated with ZO-1. The connexin of dendrites of B-type cells is not yet known. In mouse, Cx57 is expressed on both dendrites and axon terminals of the horizontal cells (Janssen-Bienhold et al., 2009). However, ZO-1 is only expressed and colocalized with Cx57 on the horizontal cell dendrites beneath the cone pedicles, and not on the axon terminals.

Schematic illustration of two photoreceptor terminals, gap junctions, and their synaptic organization. A, Vertical view of a cone pedicle and a neighboring rod spherule. Within the terminals, ribbons with tethered vesicles are depicted. Corresponding invaginating elements of postsynaptic cells are formed by dendritic tips of horizontal cells and ON bipolar cells. OFF bipolar cells making flat contacts to the cone pedicles are electrically coupled by Cx36-containing gap junctions (blue rectangles), which are also formed between photoreceptors. The red bars beneath the pedicle indicate desmosome-like junctions between horizontal cells, where ionotropic GluRs are expressed. Gap junctions are also formed between their dendrites (green rectangles) at the same level and in close vicinity to the desmosome-like junctions. B, Model of the spatial arrangement of proteins at the investigated synapses in the OPL. The gap junction is closely associated with the GluRs of desmosome-like junctions and surrounded by tight or adherens junctions formed by ZO-1 and transmembrane proteins. ZO-1 links these proteins—and most likely not the connexins—to the cytoskeleton.
Gap junction architecture
An important finding of our study is that ZO-1 is not colocalized with Cx50 at the giant gap-junctional plaques of A-type horizontal cells. It appears to form a kind of tight or adherens junction around the gap junctions instead of interacting with Cx50 directly. This arrangement might be comparable with electron microscopic observations in the cat retina (Kolb, 1977), where horizontal cell gap junctions are bounded by macula adherens junctions. Also, the organization of gap junctions bordered by tight junctions was shown in electron microscopic studies of rabbit corneal endothelial cells (Raviola et al., 1980).
ZO-1 was originally identified as a tight junction-associated protein (Stevenson et al., 1986) and it belongs to the family of membrane-associated guanylate kinase homologs (MAGUKs) (for review, see González-Mariscal et al., 2000). More recently, ZO-1 was found to codistribute with gap junctions at cell–cell contacts, and to interact with Cx31.9, Cx43, Cx45, Cx46, and Cx50 via its second PDZ domain (for review, see Giepmans, 2004) and with Cx35/36 via the first PDZ domain (Li et al., 2004; Flores et al., 2008).
Our results suggest that ZO-1 functions more like a tight junction protein rather than interacting directly with Cx50, Cx57, and Cx36: although Cx57 lacks a classical C-terminal PDZ domain-binding motif, ZO-1 and Cx57 are colocalized at the dendro-dendritic gap junctions in mouse horizontal cells. Furthermore, in Cx36−/− mice, labeling of ZO-1 was only slightly reduced, whereas the absence of Cx36 resulted in a large reduction of ZO-2 and ZONAB (ZO-1-associated nucleic acid-binding protein) (Ciolofan et al., 2006). These results suggest a direct interaction of Cx36 with ZO-2 and/or ZONAB rather than with ZO-1.
Based on our results, the model in Figure 9B describes the composition of proteins at gap junctions in close apposition to the GluRs at desmosome-like junctions. In our view, ZO-1 does not serve as a scaffolding protein for the connexins, but it forms margins around the gap junctions, most likely associated with transmembrane proteins such as occludin, cadherins, or other junction adhesion molecules (for review, see Giepmans, 2004). At these tight or adherens junctions, ZO-1 typically links the transmembrane protein to the cytoskeleton, while the binding partner itself spans the membrane and provides for the bonding of adjacent cells.
However, assuming that ZO-1 acts as a tight or adherens junction protein around gap junctions, a direct interaction of ZO-1 with the connexins at the outer rim of the gap junction is still possible because of its multiple binding sites. As mentioned above, ZO-1 binds to the connexins via its first or second PDZ domain. But the interactions with actin and occludin, for instance, occur via its GUK domain and its proline-rich C-terminal half, respectively (Fanning et al., 1998). Hence, ZO-1 could potentially bind to all three of these proteins simultaneously at those sites where the two types of junctions adjoin.
This kind of assembly could also explain the results of other studies, where a direct interaction of ZO-1 with different retinal connexins has been shown by coimmunoprecipitation and pull-down assays (Li et al., 2004, 2008). In addition, Li et al. (2008) presented data from electron microscopic (FRIL) experiments, indicating a coscaffolding of Cx45 and Cx36 by ZO-1 at bihomotypic gap junctions in the IPL. In their model of gap junctional architecture, ZO-1 intracellularly covers the whole area of the gap junction. In contrast, similar experiments in their study from 2004 show ZO-1 only at the outer rims of Cx36-containing gap junctions in the IPL, which in turn supports our model. It remains unclear whether these results describe varying modes of ZO-1 interaction with proteins at different gap junctions, and further investigation is needed to elucidate the structural composition of gap junctions and their associated proteins.
Modulation of horizontal cell coupling
Horizontal cell coupling is one mechanism that allows these cells to collect light information over a large area of the retina and to adjust the gain of photoreceptors to different levels of ambient light (for review, see Kamermans and Spekreijse, 1999). The coupling is modulated by ambient light in a triphasic manner: at low and high levels, coupling is reduced, while at intermediate light levels, coupling is maximized (Xin and Bloomfield, 1999). Various studies have shown that horizontal cell coupling is reduced by endogenous release or application of dopamine. The activation of D1 receptors leads to an increase in cAMP, which in turn leads to the phosphorylation of connexins by protein kinase A (Piccolino et al., 1984; Lasater, 1987; Hampson et al., 1994; McMahon, 1994; He at al., 2000). Another pathway for modulation of gap junction is via nitric oxide, which activates soluble guanylate cyclase, leading to an increase in cGMP and a reduction in gap-junctional conductance (Lu and McMahon, 1997; Pottek et al., 1997; Xin and Bloomfield, 2000). Both pathways lead to reduced horizontal cell coupling, but work through different mechanisms, suggesting several distinct pathways for dark- and light-induced modulation. Endogenous dopamine release is increased by light and during prolonged darkness at night (Weiler et al., 1997), but what happens at low light levels or darkness during the day?
We found two kinds of gap junctions associated with ZO-1: giant gap-junctional plaques at horizontal cell dendritic crossings in the OPL, and small gap-junctional plaques beneath the cone pedicle, where the dendrites of horizontal cells converge. The small gap junctions were in close relationship with clusters of GluRs at the desmosome-like junctions in monkey and rabbit retinas, indicating that calcium may play a role in regulating gap-junctional permeability. In darkness, horizontal cells are depolarized and calcium enters the cell via calcium-permeable AMPA receptors and voltage-gated calcium channels (Solessio and Lasater, 2002; Schubert et al., 2006). In the past, the role of calcium in coupling regulation has been discussed controversially (Spray et al., 1982; Lazrak and Peracchia, 1993), but calcium was recently found to regulate the permeability of Cx43 in a calmodulin-dependent manner (Lurtz and Louis, 2007). Therefore, calcium entering through discrete sites, such as the GluRs at desmosome-like junctions, may regulate gap-junctional coupling during conditions of retinal adaptation which are not triggered by dopamine.
A comparable system can be found at synapses formed by club endings of auditory afferents onto teleost Mauthner cells. These are mixed synapses where glutamatergic synapses work in parallel with gap junctions formed by Cx35 (for review, see Pereda et al., 2004). The conductance of these electrical synapses is regulated by dopamine and NMDA receptor activation, and different pathways are used: cAMP-dependent in the case of dopamine, and calcium-dependent in the case of NMDA receptor. Mixed synapses have also been identified in rat spinal cord and inferior olive (Rash et al., 1996; Pereda et al., 2004). Pereda et al. (2004) proposed that GluR-mediated modulation of electrical synapses is not limited to mixed synapses and can be found where chemical synapses are close to gap junctions. In contrast to the paracrine action of dopamine released from somewhat distant varicosities, modulation by GluRs is more spatially restricted, differentially controlling the conductance of nearby gap junctions. According to our data, we suggest that the giant horizontal cell gap junctions are modulated by dopamine, whereas the small dendritic gap junctions beneath the cone pedicles are modulated by calcium and possibly also by dopamine.
Footnotes
-
This study was supported by a grant of the Deutsche Forschungsgemeinschaft (FOR 701, PE 38/16-1). We thank B. Marshallsay, G.-S. Nam, and W. Hofer for excellent technical assistance, J. Nathans for providing the S-opsin antibody, and J. Trümpler for editing this manuscript. We give special thanks to H. Wässle for his continuous support and input.
- Correspondence should be addressed to Silke Haverkamp, Max-Planck-Institut für Hirnforschung, Deutschordenstrasse 46, D-60528 Frankfurt/Main, Germany. haverkamp{at}mpih-frankfurt.mpg.de





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}