

Abstract
In seasonally breeding birds, the vernal growth of the song system is thought to result primarily from increased daylength and the associated increase in circulating testosterone. Other environmental factors such as social cues between mates influence the timing of reproduction, but less is known about how social cues might affect the song system and song behavior. We used white-crowned sparrows (Zonotrichia leucophrys gambelii) to test the hypothesis that the presence of a female in breeding condition influences song nuclei and song behavior of adult males. There were four treatment groups: (1) eight males housed individually in the same room on long days and paired with estradiol-implanted females; (2) eight males housed similarly on long days but without females; (3) four males isolated on long days; and (4) four males isolated on short days. The volumes of two song nuclei, HVc and RA, were significantly larger in males housed with females than in any other treatment group. Males isolated on short days had smaller HVc, RA, and area X volumes than all other groups. The volumes of Rt (a thalamic nucleus not involved in song) and the telencephalon did not differ among groups. Plasma androgen levels did not differ among the three long-day, social treatment groups at the times sampled, but were lower in the short-day isolates. Males paired with females sang at a higher maximum rate than males housed together, who sang at a higher rate than long-day isolates. These results suggest that seasonal plasticity in the adult song system is influenced by social cues.
Song behavior is an important aspect of reproduction in birds (Catchpole and Slater, 1995; Kroodsma and Miller, 1996). Birdsong is used both to attract mates and to declare a territory from which other birds are aggressively excluded. Several features of song vary across seasons in birds that breed seasonally (Brenowitz and Kroodsma, 1996; Smith et al., 1997a; Brenowitz et al., 1998). The seasonal variability in song behavior is accompanied by structural changes in the brain regions that control song learning and production. The volumes of the neostriatal nucleus HVc, the robust nucleus of the archistriatum (RA), area X of the parolfactory lobe, and the tracheosyringial portion of the hypoglossal nucleus (nXIIts) can be as much as 188% larger in breeding birds than in nonbreeding birds (Nottebohm, 1981; Arai et al., 1989; Kirn et al., 1989; Brenowitz et al., 1991; Rucker and Cassone, 1991; Bernard and Ball, 1995; Smith et al., 1995, 1997a,b,c; Smith, 1996; Gulledge and Deviche, 1997). An increase in circulating levels of testosterone caused by increasing daylength is thought to be the primary proximate cue that induces growth of the song system and changes in song behavior (Smith et al., 1997b). Other environmental factors such as social cues between individuals are also known to influence steroid levels, the timing of reproduction, and the expression of song behavior (Bischof et al., 1981; Moore, 1983; Wingfield and Kenagy, 1991). The role that these social cues might play in the seasonal growth of the song system has, however, remained unexplored.
In several nonavian species, socially induced changes in adult behavior have been associated with cellular and molecular changes in the nervous system. Male Sprague Dawley rats housed with sexually receptive females have smaller motor neurons in the spinal nucleus of the bulbocavernosus than males housed with unreceptive females (Breedlove, 1997). In African cichlid fish (Haplochromis burtoni), changes in social status and reproductive state are accompanied by changes in the size of gonadotropin-releasing hormone cells in the ventral forebrain (Francis et al., 1993; Fernald, 1995; White and Fernald, 1997). In crayfish (Procambarus clarkii), the lateral giant tailflip circuit responds differently to serotonergic innervation depending on the social status of the animal (Yeh et al., 1996). In parthenogenetic whiptail lizards (Cnemidophorus uniparens), sociosexual behaviors of displaying individuals can influence the expression of estrogen and progesterone receptors in the hypothalamus of their cagemates (Hartman and Crews, 1996).
Given that social interactions can influence neuronal attributes in several species, we investigated whether the seasonal anatomical changes in the song system are also influenced by social cues. In this study we tested the hypothesis that social cues induce an increase in the size of song nuclei and changes in the behavior that these nuclei control. We found that HVc and RA significantly increased in volume (20 and 15%, respectively) in response to social cues. Furthermore, social cues induced a 45% increase in maximum song production rate.
MATERIALS AND METHODS
All protocols used in this experiment were approved by the University of Washington Animal Care Committee and were in accordance with the National Institutes for Health Guide for the Care and Use of Laboratory Animals.
Animal collection and housing. We collected juvenile white-crowned sparrows from eastern Washington during the autumnal migration in September 1995. These birds were ∼3–4 months old. We used juveniles so that all birds in this study would be the same age and have the same reproductive history. All birds were brought to the University of Washington and raised to adulthood (∼16–17 months of age when killed). Initially, all birds were housed together in outdoor aviaries under ambient conditions. Food and water were availablead libitum throughout the experiment. On January 23, 1996, all birds were sexed by laporotomy and placed into an environmental chamber in individual cages on a short-day photoperiod (8 hr light) at 20°C (Fig. 1A); this chamber allowed us to control photoperiod and temperature. On February 7, we photoshifted all birds overnight to a long-day photoperiod typical of what they experience on their Alaskan breeding grounds (20 hr light). We maintained them on this photoperiod until April when they molted into adult plumage. Once the molt was complete in late May, all birds were gradually photoshifted down (minus 1–2 hr per day) to short days, where they remained for 12 weeks to ensure photosensitivity.
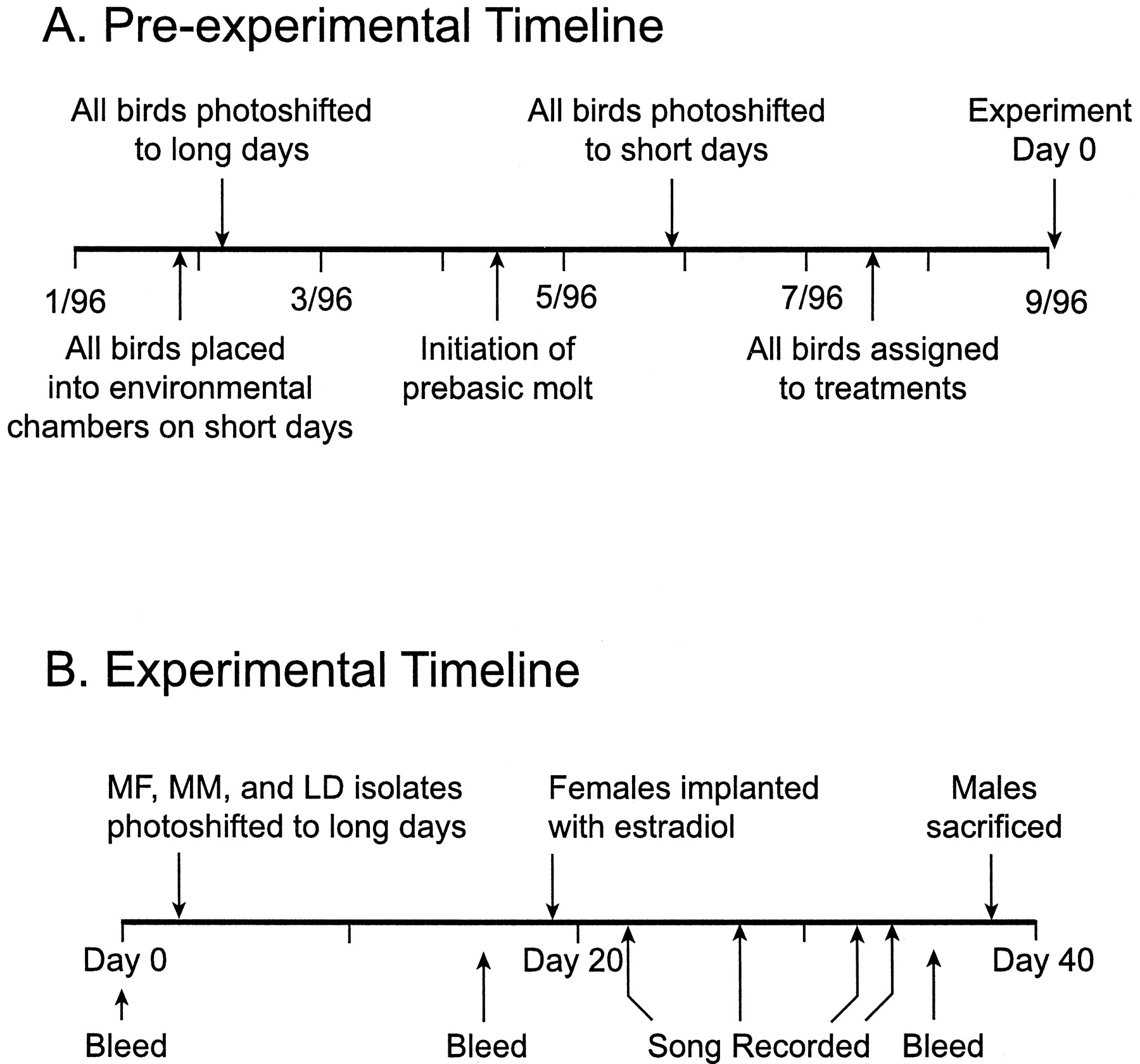
Pre-experimental (A) and experimental (B) timelines showing when birds were photoshifted, bled, recorded, and killed.
Experimental design/timeline. All birds were randomly assigned to one of four treatments. Eight males were housed in individual cages in the same room on long days. Each of these males shared his cage with a female implanted with estradiol (E2) to stimulate breeding behavior. Wild-caught females will not perform breeding behaviors in captivity unless they are implanted with E2 (Moore, 1983). (One male from this group was excluded from all analyses because his female cagemate lost her estrogen implant during the study.) Birds in this room were in visual and acoustic contact with every other bird in the room. This group will be referred to as the “MF” treatment throughout the remainder of this paper. Eight males were housed identically to those in the MF group, but without females (male–male or MM group). We housed four males in total isolation from each other in acoustic attenuation boxes on long days (LD isolates). Four additional males were housed identically to the LD isolates but on a short-day photoperiod (SD isolates).
On September 3, 1996 (day 2 of the experiment), birds in the long-day treatment groups were photoshifted overnight to 20 hr daylengths (Fig.1B). Seventeen days later, females were implanted subcutaneously with 10 mm of E2 in SILASTIC tubing (1.47 mm inner diameter × 1.96 mm outer diameter) to induce plasma concentrations of this hormone typical of wild-breeding birds. We chose to augment estrogen in females at this time (instead of day 2) to allow endogenous hormone levels to rise gradually. After implant, all females expressed E2-dependent behaviors (chitter vocalizations, copulation solicitation, and copulation). Blood samples for hormone analysis were collected from all males on days 0, 16, and 37. Songs were recorded on days 22, 27, 32, and 34. Males were killed between days 38 and 39, and their brains were removed for histological analysis (Fig. 1B).
Histology. Male sparrows were deeply anesthetized by methoxyflurane inhalation and killed by transcardial perfusion with 0.9% saline followed by 10% neutral buffered formalin. Brains were removed and stored in fixative for at least 4 weeks; post-fixation duration was balanced across groups. After completion of post-fixation, brains were embedded in gelatin and cryoprotected in a 20% sucrose/formalin solution for 3–4 d. Transverse frozen sections were cut at 50 μm on a freezing microtome and collected into saline. Alternate sections were mounted onto gelatin-coated slides and Nissl-stained with thionin. In white-crowned sparrows and other species, the Nissl-defined borders of song nuclei coincide with the borders as defined by other labels (Johnson and Bottjer, 1993; Bernard and Ball, 1995; Smith et al., 1997c; Soma et al., 1997; Tramontin et al., 1998).
Morphometry. We measured the volumes of four telencephalic song nuclei: HVc, RA, lMAN, and area X. We also measured the volume of the rotund nucleus (Rt), a thalamic nucleus not involved in song control. Our measurements of HVc included the caudomedial extension referred to as para-HVc by Johnson and Bottjer (1995) and therefore coincide with the measurements of Smith et al. (1995, 1997c). We projected an image of each section onto paper at final magnification of 46×. We traced the Nissl-defined outline of each brain nucleus profile, scanned the tracings into a microcomputer, and calculated the area of each nucleus profile using NIH Image software (version 1.57; Wayne Rasband, National Institutes of Health). We estimated the volume of each nucleus using the formula for a cone frustum over each measured area (Smith et al., 1995, 1997b).
We also measured the volume of the entire telencephalon in which the song nuclei reside. In each brain, we traced either the right or left telencephalon (alternated systematically after a random start). We defined the borders of the telencephalon as in DeVoogd et al. (1993)and Brenowitz et al. (1998). In the rostral and caudal extents of the telencephalon, we traced the entire telencephalic lobe. In sections where the telencephalon was contiguous with the diencephalon, the septomesencephalic tract, anterior commisure, and occipitomesencephalic tract were used as natural borders of the telencephalon. We projected an image of every third section of the telencephalic hemisphere (300 μm sampling interval) onto paper at a final magnification of 14×, traced the borders, and scanned these tracings into a microcomputer. Telencephalon hemisphere volume was estimated with the formula for a cone frustum and multiplied by 2 to obtain total telencephalon volume; no differences were detected between the left and right telencephalic lobes. All brain regions were traced blind to the treatment assignment of each bird.
To control for possible differences in body size, overall brain size, or histological preparation, we calculated the volumes of HVc, RA, area X, and lMan relative to either Rt volume or total telencephalon volume. As a further control for differences in telencephalon volume among groups, we performed a residual analysis (DeVoogd et al., 1993;Brenowitz et al., 1998). Briefly, we log-transformed brain nuclei volumes and plotted the values for all four treatment groups against the log-transformed telencephalon volume minus the volume of the nucleus under scrutiny. After fitting a single regression line through the plot, residuals were recorded and compared among groups with a one-way ANOVA.
Behavioral analyses. We recorded singing behavior from all treatment groups twice per day on four different days (Fig.1B) (total = eight sessions per treatment group). One recording session was always at 8:00 A.M. (3 hr after lights on). The second session time was randomized to control for a possible diel rhythm in singing behavior. The tape recorder ran continuously during each 30 min recording session. We used Sony TCD5M recorders and Sennheiser ME 80 directional microphones for the MF and MM treatments and Realistic tie-clip microphones for LD and SD isolates. We recorded multiple birds onto single tapes in the MF and MM groups, but each bird had a unique song type that allowed identification of who was singing during the session. In each treatment group, we measured the average song rate for each recording session and maximum sampled song rate for each individual bird. Maximum sampled song rate was obtained by scanning through all of our recordings and finding the 5 min during which each bird produced the most song. We chose a 5 min assay interval because song bout lengths were variable in these birds but always lasted at least 5 min. SD isolates never sang and so were not included in any statistical analyses of song production.
We measured song stereotypy in two treatment groups (MF and MM). LD isolates sang too infrequently to allow statistical analysis of song stereotypy, whereas SD isolates never sang throughout the entire experiment. To obtain high-quality recordings for stereotypy analysis, tie-clip microphones were attached to each cage so that recordings of specific males’ songs could be made. These recordings were ∼20 min in duration. Ten songs from each male were analyzed to measure song stereotypy using customized software (J. Burt, University of Washington). We digitized each song and displayed it on the computer screen as a sound spectrogram. On this spectrogram, we used time and frequency cursors to measure temporal and spectral attributes of song. As measures of song stereotypy, we collected data on 14 different song attributes (Fig. 2, see Table 3). To determine whether groups differed in absolute values of song attributes, we compared means between groups. To determine whether groups differed in the variability of these attributes, we compared coefficients of variation (CV = SD/mean × 100) between the MF and MM treatment groups. Song attributes were measured blind to treatment group.

Sound spectrogram showing terminology used to describe Gambel’s white-crowned sparrow song.
Hormone assays. Blood samples were taken from all birds at three different times (Fig. 1B). We collected 300 μl of whole blood by alar venepuncture into heparinized capillary tubes. The blood was immediately centrifuged, and the plasma was removed and stored at −20°C until assay.
Plasma androgen levels were measured in a single radioimmunoassay (RIA) using the method of Ball and Wingfield (1987), except that we did not use column chromatography to separate testosterone (T) from dihydrotestosterone (DHT). The T antiserum we used had a 60% cross reactivity with DHT, so all results are conservatively stated as plasma concentration of androgens. DHT is present in much lower levels than T in the plasma of white-crowned sparrows and parallels T throughout the breeding cycle (Wingfield and Farner, 1978).
Plasma samples (100 μl) were allowed to equilibrate overnight with 2000 cpm of tritiated T in 300 μl of distilled H2O. Androgens were extracted in 5 ml of dichloromethane, dried, and reconstituted in 550 μl of PBS with 2% gelatin and 2% sodium azide. Samples were vortexed and refrigerated overnight at 4°C. We removed 100 μl of sample into a vial with scintillant that was counted on a scintillation counter and compared with a standard for determination of percent recovery of steroid after extraction. We processed duplicate 200 μl aliquots of each steroid sample for RIA by adding 104 cpm of tritiated T and antiserum to T (Wien Laboratories). In parallel, we processed a standard curve consisting of two duplicate sets of tubes containing known amounts of unlabeled T. After overnight incubation, androgens bound to antibodies were separated from free androgen with dextran-coated charcoal. Bound steroid was decanted, added to scintillant, and counted on a Beckman scintillation counter. The minimum detectable concentration of androgen ranged between 0.07 and 0.11 ng/ml depending on the plasma sample volume. Samples with undetectable levels of steroid were scored at the limit of detection.
Plasma luteinizing hormone (LH) was measured in a single RIA using the method of Follett et al. (1972, 1975). Duplicate aliquots of plasma (10–20 μl each) were processed in parallel with a triplicate series of tubes containing known concentrations of LH. LH antiserum (Sharp et al., 1987) in normal rabbit serum was added to all tubes and allowed to incubate for 24 hr. [125I]LH was then added and allowed to incubate for 24 hr. Goat anti-rabbit precipitating serum was added and allowed to incubate for 24 hr. Finally, samples were diluted, centrifuged, and aspirated. The resulting dry pellets containing the bound LH were counted on a Beckman 5500 gamma counter. The minimum detectable limit for the assay was 0.039 ng/ml. No samples were below this detectable limit.
Statistics. Comparisons between groups were done with one-way ANOVAs. Post hoc pairwise comparisons were performed using Fisher’s protected least significant difference tests (PLSD). Planned comparisons were performed on hormone data using PLSD tests. Song stereotypy was compared between the MF and MM treatment groups using Student’s t tests. The α level for all statistical comparisons was 0.05 (two-tailed).
RESULTS
Brain morphometry
HVc and RA were both significantly affected by treatment (F(3,21) = 24.43, p < 0.001;F(3,22) = 26.40, p < 0.001, respectively), and were larger in the MF males than in any other group (p < 0.05 for all comparisons; PLSD) (Fig.3A). HVc and RA did not differ between the MM group and the LD isolates (p > 0.05 for both comparisons; PLSD), but both groups had larger HVc and RA volumes than the SD isolates (p < 0.05 for all comparisons; PLSD). Area X was also significantly affected by treatment (F(3,22) = 9.57; p < 0.001). Area X did not differ among the three long-day treatment groups, but was significantly smaller in the SD isolates (p< 0.01; PLSD) (Fig. 3B). lMAN volume did not differ between groups (F(3,22) = 2.52; p = 0.089) (Fig. 3B).
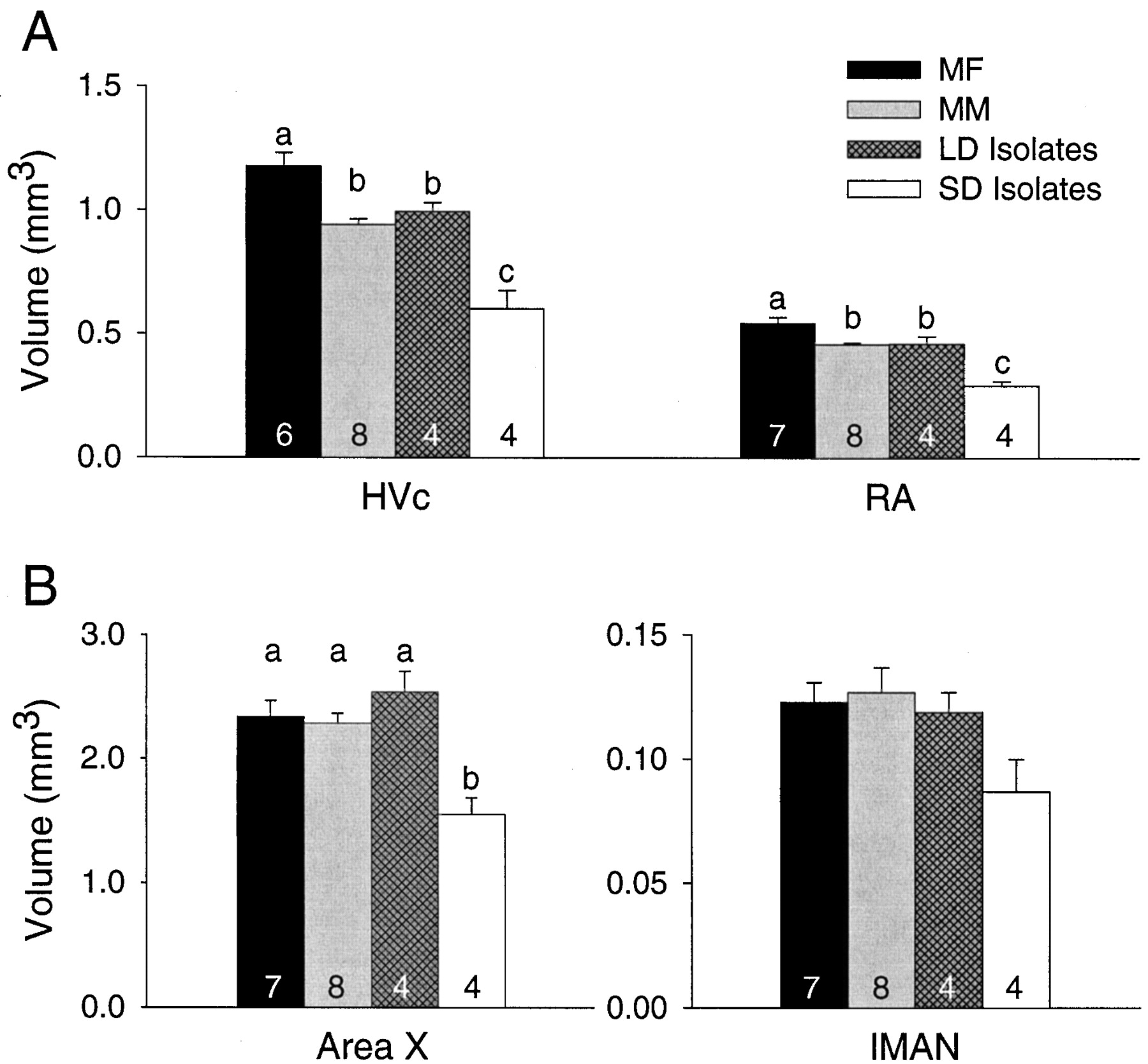
Volumes of four telencephalic song control nuclei in either (A) the main descending motor pathway (HVc, RA) or (B) the anterior forebrain pathway (IMAN, Area X). HVc and RA were influenced by social and photoperiodic cues. Area X was only influenced by photoperiodic cues. MF, Males housed with females; MM, males housed with other males;LD Isolates, males isolated on long days; SD Isolates, males isolated on short days. Numbers within bars are sample sizes, and letters above barsrepresent statistically different groups (PLSD; p< 0.05).
Neither Rt volume nor telencephalon volume differed among groups (Table1). When we normalized song nuclei volumes by dividing them by Rt volumes, the differences between groups persisted as described above. Song nuclei volumes expressed relative to telencephalon volume differed among groups as described above, but with one exception. HVc volume relative to telencephalon was still largest in the MF group, which differed significantly from the MM group and the SD isolates. We did not, however, detect a significant difference between the MF group and the LD isolates. This was likely caused by the small sample size in the LD isolate group (power of test, 0.149). The residual analysis yielded the same results as did the analysis of song nuclei volumes relative to telencephalon volume (Table2).
Brain morphometry
Analysis of residuals for volumes of brain nuclei relative to telencephalon volume
Behavioral analysis
Average song rate in each recording session did not differ between groups as a function of treatment or time (F(2,15) = 2.04, p = 0.164;F(7,105) = 0.17, p = 0.990). The interaction between treatment and time was also not significant (F(14,105) = 0.57; p = 0.886). We did find significant differences between groups in maximum sampled song production rate (F(2,18) = 15.64;p < 0.001) (Fig. 4). The maximum sampled song rate recorded from birds in the MF treatment groups was significantly greater than the maximum rate of both the MM group (p < 0.010; PLSD) and the LD isolates (p < 0.010; PLSD). Maximum sampled song rate in the MM birds was significantly greater than that in the LD isolates (p < 0.050; PLSD), but this result should be viewed with caution because only two of the four LD isolates ever sang. Song stereotypy parameters did not differ between MF and MM treatment groups (Fig. 2, Table 3).
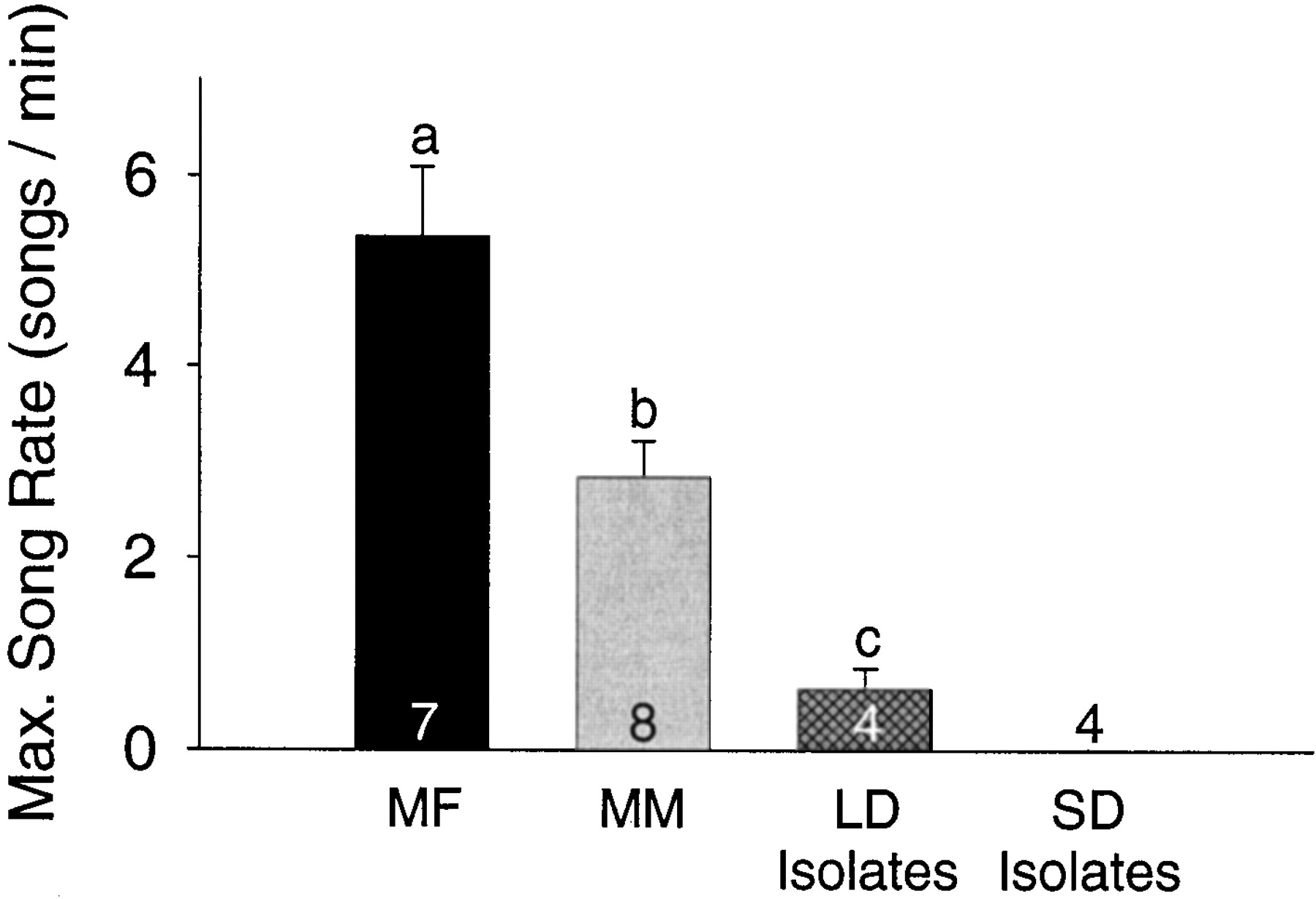
Maximum song rate differed significantly among the three long-day groups. The SD isolates are included in this graph, but did not sing and so were not included in the statistical analysis.MF, Males housed with females; MM, males housed with other males; LD Isolates, males isolated on long days; SD Isolates, males isolated on short days.Numbers within bars are sample sizes, and letters above bars represent statistically different groups (PLSD;p < 0.05).
Behavioral song attributes
Hormone analysis
Plasma androgen levels were significantly affected by treatment and by time with no interaction between these two factors (F(3,18) = 3.09, p = 0.05;F(2,36) = 9.01, p = 0.001; andF(6,36) = 1.68, p = 0.154, respectively) (Fig. 5). Planned comparisons revealed a difference in androgen levels between the MM group and the SD isolates at day 17 just before the females in the MF group were implanted with E2 (p < 0.05; PLSD). On day 36 (just before kill), the SD isolates had significantly less plasma androgen than did the MF, MM, or the LD isolates (p < 0.001; p < 0.05; and p < 0.001, respectively; PLSD). The MF, MM, and LD isolates did not differ from one another at any sampling time (allp values > 0.05; PLSD).

Plasma androgens (top) and luteinizing hormone (bottom) did not differ between social treatment groups. Both androgens and LH were, however, affected by photoperiod. Asterisks represent statistically significant differences between treatments (PLSD; p< 0.05).
Plasma levels of LH were significantly affected by treatment and by time, with a significant interaction between treatment and time (F(3,17) = 34.06, p < 0.001;F(2,34) = 20.51, p < 0.001; andF(6,34) = 3.39, p = 0.010, respectively) (Fig. 5). Planned comparisons revealed an effect of treatment at 17 and 36 d after photoshift. At both times, the SD isolates had significantly lower levels of plasma LH than did all other treatment groups (PLSD; p < 0.001 for all three pairwise comparisons).
DISCUSSION
The presence of females in breeding condition significantly increased the volumes of HVc and RA in their adult male cagemates. This increase occurred in the absence of any changes in Rt or telencephalon volume. Furthermore, no other song nuclei measured showed any response to social stimulation. These data suggest that the influence of social cues was specific to the main descending motor pathway within the song system that includes HVc and RA. These volume differences between treatment groups might represent differences in the magnitude of HVc and RA growth, or they might reflect a shift in the time course of photoperiodically induced song system growth. Our data cannot discern between these two possibilities.
The source of these stimulatory social cues is unclear. These cues might have come directly from the female cagemate, or they may have resulted from aggressive song and/or visual interactions between males that are associated with mate guarding (Moore, 1984; Wingfield, 1988). These alternative hypotheses might be distinguished by isolating male–female pairs and comparing them with pairs that are in visual and acoustic contact with other pairs. Another goal for future studies will be to determine which specific behavioral cues induce growth in HVc and RA. A study conducted by Moore (1983) provides suggestions as to the nature of these behavioral cues. He found that males caged with E2-implanted females had higher levels of LH and T than males housed with blank-implanted females. In captivity, only females treated with E2 will perform breeding behaviors such as copulation solicitation, copulation, and chitter vocalizations. Moore concluded that it was these E2-dependent female behaviors that stimulated increased T and LH levels in male cagemates. These same behavioral cues might be directly or indirectly responsible for the growth seen in HVc and RA.
Increased photoperiod alone induced growth of HVc, RA, and area X in the MM group and in the LD isolate group in our study. This result is consistent with data on free-living white-crowned sparrows (Soma et al., 1997; Brenowitz et al., 1998), except that the magnitude of the photoperiodically induced song nucleus growth is slightly smaller than that reported by Soma et al. (1997) in wild Gambel’s white-crowned sparrows. Smith et al. (1995) were unable to induce changes in the song system of captive Gambel’s white-crowned sparrows with increased daylength alone. Our results may differ from those of Smith et al. (1995) because we used young birds with no reproductive experience in the field, whereas they used birds captured as adults. Such experience may influence an individual’s response to a single environmental cue such as photoperiod in the laboratory.
The presence of females stimulated a pronounced change in male song behavior. Males paired with females expressed a higher maximum song rate than males without females. Evidence from several species indicates that in the context of mate choice, females prefer males that sing at higher rates (Radesater et al., 1987; Wasserman and Cigliano, 1991; Houtman, 1992; Collins et al., 1994; Searcy and Yasukawa, 1996). High song rates are also associated with aggression and dominance between males (Wingfield and Hahn, 1994; Otter et al., 1997; Bower, 1998).
The socially induced increase in maximum song rate occurred in the absence of any differences in average song rate. Males in the MF group could sing at a higher rate than MM males, but did not do so consistently throughout the experiment. As a result, males in both treatments were exposed to similar overall amounts of singing but differed with respect to song system anatomy. Two of the four LD isolates were never observed singing but had song nuclei of similar size to LD isolates who did sing and to males in the MM group. This observation might suggest that seasonal changes in song nucleus volume can occur in the absence of either song production or hearing song. A strong test of this hypothesis will require more extensive sampling of song behavior and direct manipulations of song production and perception.
The greater maximum song rates and large premotor song nuclei in the MF birds might imply the existence of neural constraints on song rate. This observation might be consistent with data from two different morphs of white-throated sparrows (Zonotrichia albicollis) that differ in song rate and in the anatomy of some song-related nuclei (DeVoogd et al., 1995). It is problematic, however, that white-throated sparrow morphs significantly differ in the sizes of area X and lMAN, which are not required for production of adult song (Bottjer et al., 1984; Sohrabji et al., 1990; Scharff and Nottebohm, 1991), but not in HVc and RA, which are necessary for song production and where we see socially induced volume changes. Data from European starlings (Sturnus vulgaris) suggest that individual differences in HVc and RA correlate with differences in song bout length, but the relationship between song nucleus size and song rate remains unclear (Bernard et al., 1996). Furthermore, the volumes of HVc and RA are not correlated with song rate in eastern marsh wrens (Cistothorus palustris) (Brenowitz and Kroodsma, 1998). Our data need not indicate that increased maximum song rate is causally related to increases in HVc and RA volume. Differential motivational levels between groups could also have accounted for the differences in song production rate.
Plasma androgen levels at the times we measured them responded to changes in photoperiod but not to social manipulation. Long photoperiods induced increases in plasma levels of androgens and LH. The mean androgen levels in our long-day birds were ∼50% of those seen in wild-breeding white-crowned sparrows (Wingfield and Farner, 1978). Nonetheless, it appears that these levels were sufficient to cause changes in secondary sex characteristics (cloacal protuberance length; data not shown) and song system anatomy. We did not detect any social influence on plasma levels of either of these hormones. These data can be interpreted in several ways.
One possibility is that the mechanism controlling socially induced changes in song system anatomy are steroid-dependent, but plasma levels of steroids do not accurately reflect local concentrations in the brain. Gonadal steroids and steroid precursors can be converted to their active forms by enzymes located in the brain (Vanson et al., 1996; Schlinger, 1997). We cannot rule out the possibility that in the brain, local concentrations of T (or any of its active metabolites including DHT and E2) might have differed between our groups. This issue might be resolved by performing RIA on brain tissue homogenates.
Another possible interpretation of our data are that plasma concentrations of androgens differed between social treatments but we failed to detect these differences. As mentioned earlier, Moore (1983)found that white-crowned sparrow males paired with E2-implanted females had significantly higher levels of T and LH than males housed with blank-implanted females. Moore took blood samples five times in the 40 d after the photoshift to long days, whereas we took only three samples over a similar number of days. We sampled less often in an attempt to minimize stress that might affect the brain morphology and song behavior. With our reduced sampling, it is possible that we missed a hormone peak that may have revealed differences between social treatments.
A third interpretation of our data is that social influences do not affect changes in the brain through steroid-dependent mechanisms. We consider this unlikely in light of the social effects on plasma T uncovered by Moore (1983), and given that seasonal changes in the song system are correlated with changes in plasma T (Smith et al., 1995,1997a,b; Brenowitz et al., 1998). Our data, however, cannot rule out this third hypothesis.
In summary, we have shown that social cues can enhance the seasonal growth of the motor pathway of the adult song system and can induce changes in the behavior that this circuit controls. This is the first indication that environmental cues other than photoperiod can influence the anatomy of the song control system. This reinforces the need to consider social and other environmental cues in designing laboratory studies of brain and behavior. There are many other examples of environmental cues that animals use to time reproduction, such as temperature, precipitation, and food availability (Wingfield and Kenagy, 1991). Perhaps the seasonal plasticity in the song system responds to an array of cues from an individual’s surroundings.
Songbirds display the most dramatic socially induced neuronal reorganization in the forebrain of any vertebrate studied thus far. Although the song system may provide an excellent model for studying the effects of social cues on the adult brain, songbirds are not unique in their sensitivity to social stimulation. Social cues are emerging as important influences on brain anatomy and function across a variety of taxa (Fernald, 1995; Yeh et al., 1996; Breedlove, 1997; White and Fernald, 1997). The effects of these cues on the song system may be analogous to the effects of environmental enrichment on cortical structure in rats (Bennett et al., 1964; Green et al., 1983) and hippocampal neurogenesis in adult mice (Kempermann et al., 1998). The laboratory is impoverished of the many environmental cues on which birds depend for successful reproduction and, perhaps, song system growth. The stimuli provided by females and competitors in the laboratory in this study might more closely approximate the array of cues available in a bird’s natural environment. The mechanisms by which social stimulation influences neural structure and function, however, remain to be elucidated.
Footnotes
This work was supported by National Institutes of Health Grant MH53032 to E.A.B and National Science Foundation (NSF) Grant IBN-9631350 to J.C.W. A.D.T. is supported by the NSF. We thank Karin Lent for assistance with histology, and Lynn Erckmann and Renee Crain for assistance with hormone assays. Troy Smith provided thoughtful suggestions regarding the design of this study. Kiran Soma and anonymous reviewers provided comments on this manuscript.
Correspondence should be addressed to Anthony D. Tramontin, University of Washington, Department of Zoology, Box 351800, Seattle, WA 98195-1800.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}